Ottica Campagnacci
Ottica Campagnacci










Fabrizio Campagnacci

www.otticacampagnacci.com
/home/blog

OTTICA CAMPAGNACCI aderisce alla raccolta degli occhiali usati:





OTTICA
CAMPAGNACCI
is authorized dealer of:

- Binocoli e cannocchiali
- Microscopi
- Strumenti elettronici meteo
- Altimetri e bussole
- Barometri, termometri e Galileiani
- Lenti d'ingrandimento
- Telescopi
- Visori notturni "Yukon"
- Strumenti "Suunto"
- Catalogo "C.A.I."



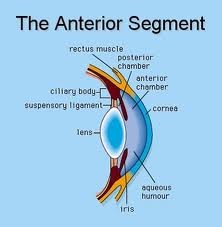
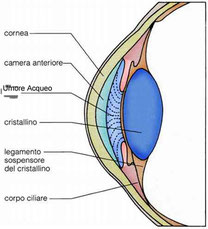
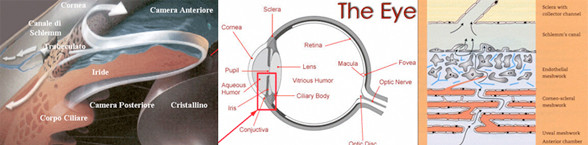
Camera anteriore e posteriore, umore acqueo


Camera posteriore
La camera posteriore, compresa tra l'iride e le formazioni che vincolano il cristallino al corpo ciliare, è più piccola e funge da bacino di raccolta iniziale dell'umore acqueo prodotto dai processi ciliari. L'umore acqueo, scorrendo lentamente tra la faccia posteriore dell'iride e quella anteriore del cristallino, si trasferisce dalla camera posteriore in quella anteriore, da dove viene drenato attraverso il sistema trabecolare.
La zona di produzione dell'umor acqueo è costituita dai processi ciliari. Sono circa 70, costituiti da creste lamellari inserite nella faccia interna del corpo ciliare. Nella struttura dei processi ciliari si distingue l'epitelio e lo stroma.
Epitelio ciliare
E' suddiviso in due strati monocellulari (interno ed esterno) affrontati reciprocamente per la parte apicale. Si comporta come
un'unità funzionale e viene chiamato sincizio epiteliale ciliare.
Strato interno o epitelio non pigmentato. Contiguo alla camera posteriore, dalla quale lo divide una membrana basale (membrana limitante interna), è costituito da cellule chiare.
Tali cellule presentano un nucleo ovale, numerosi mitocondri, apparati di Golgi, lisosomi. In particolare sono dotate di una propria membrana di rivestimento provvista di giunzioni con le cellule
contigue e, soprattutto, di numerose invaginazioni, profonde ed irregolari (betacitomembrane) che provocano un aumento della sua superficie.
Strato esterno o epitelio pigmentato. Contiguo allo stroma ciliare, è costituito da cellule pigmentate, di aspetto pavimentoso. Il pigmento, in granuli, è presente soprattutto
nella parte esterna del protoplasma.
Stroma ciliare
E' composto da tessuto connettivo contenente fibre collagene, fibre elastiche, elementi cellulari (melanociti e fibrociti) e da una rete vascolare assai sviluppata, soprattutto in prossimità dello strato epiteliale dove dà luogo ad una fittissima trama di capillari di considerevole superficie. Questi vasi tuttavia non entrano in contatto diretto con l'epitelio ciliare: tra le due formazioni cellulari si interpone infatti una tenue struttura connettivale provvista di fibre nervose con funzione vasomotoria e secretoria.

Umor acqueo
L’umor acqueo è un liquido trasparente contenuto nella camera anteriore e in quella posteriore dell’occhio. Esso si estende dunque dalla cornea, attraverso il
foro pupillare, fino al cristallino, che circonda per giungere a contatto con il corpo vitreo. Ha un peso specifico di 1006, ph 7,3 e una pressione osmotica superiore a quella del sangue. Certi
elettroliti (anioni) si trovano nell’umore acqueo ad una concentrazione nettamente superiore a quella del plasma del sangue; le proteine vi sono invece presenti in quantità assai più bassa e così
pure il glucosio. Vi si trovano rarissimi elementi figurati del sangue, per lo più linfociti che aumentano considerevolmente durante i processi infiammatori dell’occhio.
La produzione dell’umore acqueo si compie, in primo luogo, per secrezione delle cellule epiteliali dei processi ciliari e inoltre per fenomeni osmotici di diffusione attraverso i vasi
sanguigni del corpo ciliare. Il liquido si riversa nella camera posteriore dell’occhio e, attraverso il foro pupillare, passa nella camera anteriore; di qui penetra nel sistema
trabecolare sclerocorneale, filtra nel canale di Schlemm e prende quindi la via delle vene episclerali e ciliari anteriori.
Il ritmo di produzione e di riassorbimento dell’umore acqueo si compie normalmente in modo tale da determinare nell’interno dell’occhio una pressione costante, pari a 18-20 mmHg. La sua
pressione aumenta nel glaucoma.
Esso esplica diverse funzioni. Innanzitutto una funzione ottica, in quanto è uno dei mezzi refrattivi dell'occhio in particolare costituisce il secondo mezzo trasparente del
globo oculare; poi una funzione statica, poiché contribuisce a contenere la pressione intraoculare; ed infine una funzione nutritiva, in quanto fornisce numerosi
nutrienti agli elementi oculari che bagna, in particolare cristallino e cornea.
Umor acqueo: caratteristiche
fisico-chimiche
|
Peso specifico
|
1,008-1,009
|
|
Indice di refrazione
|
1,337
|
|
Acqua (%)
|
98-98,5
|
|
Cloruro di Sodio (%)
|
1-1,3
|
|
Albumine ed altro
|
tracce
|
|
Pressione (mmHg)
|
14 - 20
|

Produzione dell'umor acqueo
L'umore acqueo viene prodotto alla velocità di 2,2 +/- 0,37 mm3/min. Come è noto, il movimento di fluidi attraverso le membrane biologiche avviene per meccanismi fisici (trasporto passivo), per
meccanismi biochimici (trasporto attivo) o per meccanismi combinati.
Meccanismi passivi. Non incidono per più del 25% della produzione dell'umore acqueo. Sono rappresentati dalla diffusione, la dialisi e l'ultrafiltrazione. Le ultime due rendono conto
della differenza proteica tra plasma e acqueo.
Meccanismi attivi. Il loro intervento spiega la maggiore concentrazione nell'acqueo di sostanze come sodio, cloro, acido ascorbico, acido lattico e la differenza di potenziale esistente
tra stroma ed epitelio.
La secrezione dell'acqueo dipende da un trsporto unidirezionale di soluti, che sono seguiti passivamente dall'acqua per un gradiente di concentrazione osmotica.
La secrezione unidirezionale dell'acqueo in camera posteriore (in direzione stroma-PE-NPE-camera posteriore) avviene per via transcellulare e paracellulare. Esiste un riassorbimento parziale
dell'acqueo che dalla camera posteriore ritorna alle cellule del NPE.
L'anidrasi carbonica catabolizza la reazione tra anidride carbonica ed acqua, portando alla formazione di bicarbonato e ione idogeno. Lo ione idrogeno viene scambiato con il sodio il quale però
viene immediatamente espulso ad opera della sodio/potassio fosfatasi. L'alta concentrazione di sodio e di bibarbonato (quest'ultimo espulso mediante scambiatori con il cloro e canali per la
diffusione facilitata per il bicarbonato) negli spazi intercellulari provoca lo spostamento di acqua per gradienti di concentrazione osmotica. L' inibizione dell'anidrasi carbonica o della
sodio/potassio fosfatasi (rispettivamente con acetazolamide e digossina) provoca una diminuzione della produzione di umore acqueo del 50%.

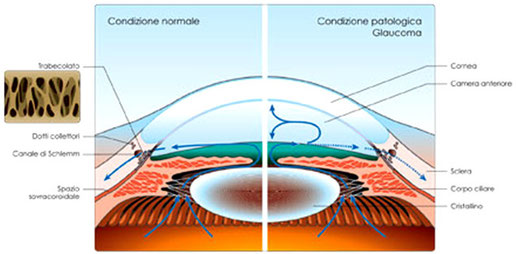
Camera anteriore
La camera anteriore è lo spazio intraoculare delimitato anteriormente dalla faccia posteriore della cornea e, posteriormente, dalla
faccia anteriore dell'iride, comunica inoltre con la camera posteriore attraverso il foro della pupilla.
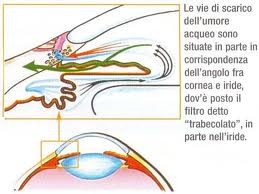
La camera
anteriore è sede di un importantissimo evento fisiologico: nel punto dove l'iride tocca la cornea, anatomicamente viene a crearsi
un angolo attraverso il cui trabecolato (canale di Schlemm) avviene il drenaggio dell'umor acqueo, liquido
fisiologico secreto dai processi ciliari e riversato in camera posteriore. L'esatto equilibrio tra prodotto e deflusso dell'umor acqueo fa si che la pressione intraoculare
(tono) esercitata dall'umor acqueo verso l'esterno si tenga costante.
Quando, per difetto di smaltimento o (più raramente) per eccesso di produzione, aumenta la quantità di umor acqueo in camera anteriore, aumenta
inevitabilmente anche la pressione ad esso associata (ipertono), la persistenza di una tale situazione potrebbe configurarsi all'interno di un quadro patologico
come il glaucoma.
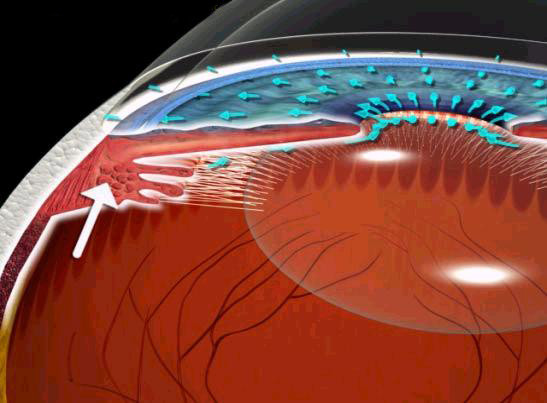
Deflusso dell'umor acqueo
L'umore acqueo, secreto direttamente in camera posteriore dai processi ciliari, attraverso lo spazio iridolenticolare passa in camera anteriore. Qui, oltre ad un rimescolio legato alle
contrazioni dei muscoli ciliari ed iridei, l'acqueo presenta regolari movimenti da parete a parete della cavità camerulare e moti circolatori di convenzione termica, ascendenti davanti all'iride
e discendenti verso la cornea, originati dalla differenza di temperatura delle due strutture.
L'attraversamento del trabecolato uveale avviene molto agevolmente grazie all'ampiezza dei pori a questo livello. L'acqueo può seguire due vie di deflusso.
Deflusso trabecolare (deflusso pressione-dipendente). Costituisce la via principale del deflusso (85%). All'altezza del trabecolato
corneo-sclerale il passaggio del liquido è meno agevole per il progressivo restringimento dei pori. A questo livello possono inoltre depositarsi detriti veicolati dallo stesso liquido. Il
trabecolato cribriforme è ancora meno permeabile: l'acqueo infatti lo attraversa solo dopo aver imbibito lo strato endoteliale e le lamelle circostanti. La sua progressione può essere accellerata
dalla contrazione delle fibre longitudinali del muiscolo di Brucke che determina l'arretramento dello sperone sclerale e l'apertura a ventaglio delle fibre trabecolari. Superato il trabecolato
cribriforme, l'acqueo deve sormontare un ulteriore ostacolo: la parete interna del canale di Schlemm che potrebbe essere, insieme al trabecolato cribriforme, la sede principale della fisiologica
resistenza al deflusso.
Deflusso uveosclerale (deflusso pressione-indipendente). Attraversato il trabecolato uveale l'acqueo abbandona la via classica per deviare
lateralmente, lungo le lamelle trabecolari, fino a giungere al tessuto uveale che riveste la faccia esterna del muscolo ciliare. Constituisce il 15% del deflusso dell'acqueo in condizioni
normali, ma potrebbe acquistare notevole importanza nelle situazioni di alterato deflusso trabecolare.
Deflusso irideo. Poco rilevante, si verificherebbe per assorbimento diretto da parte dell'iride.
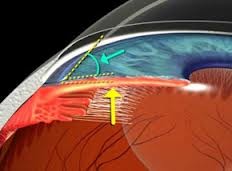
Angolo camerulare
Corrisponde al recesso periferico della camera anteriore. La parete anteriore è costituita dalla periferia della Descemet, contrassegnata da un rilievo anulare grigio biancastro (linea di Schwalbe) fatto di fibre collagene ed elastiche, e dal trabecolato attraverso il quale traspaiono il canale di Schemm e lo sperone sclerale. La parete posteriore è costituita essenzialmente dalla radice dell'iride.
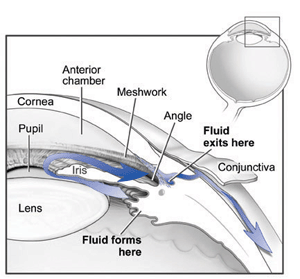
Trabecolato
Il trabecolato è una struttura deputata al deflusso dell'acqueo; nella sua porzione filtrante possiamo distinguere, dal punto di vista strutturale e funzionale, tre parti:
trabecolato uveale, trabecolato corneosclerale, trabecolato cribriforme. Il primo, quello più interno, è composto di tralci e lamelle irregolarmente disposti, ma orientati in senso radiale.
Questi tralci delimitano spazi ampi fino a 70 micron.
Il trabecolato corneosclerale è costituito da una fitta rete di lamelle che delimitano spazi intertrabecolari più ristretti rispetto a quelli dello strato uveale. A livello del trabecolato
cribriforme gli spazi tra le fibrille sono non più grandi di 4-7 micron. La struttura trabecolare contrae rapporti con i tendini anteriori dei muscoli ciliari.
Questa struttura lamellare, composta di spazi sempre più piccoli, ha caratteristiche autosimili; man mano che ci spostiamo dal trabecolato uveale a quello cribriforme la struttura resta
simile, ma cambia la scala. Il trabecolato ha tutte le caratteristiche di un frattale, tipo quelle forme solide con dimensione superiore a 2 e inferiore a 3, come la spugna di
Menger.
Certamente il trabecolato non presenta la stessa regolarità dei frattali realizzati linearmente, perché la struttura è prodotta in modo aleatorio e l'autosomiglianza è di tipo
statistico.
Perché questa struttura frattale a livello dell'organo adibito al deflusso dell'acqueo? Per permettere che una determinata struttura resista a sollecitazioni della pressione idrostatica e
alle trazioni muscolari, è necessario che la risultante degli sforzi in ogni punto non superi il limite di deformazione elastica. D'altra parte il trabecolato, essendo una struttura necessaria
alla filtrazione ed al riassorbimento dell'acqueo, deve avere una bassa densità per mantenere gli spazi sufficientemente aperti, pur mantenendo lo stesso volume e robustezza.
Per realizzare una struttura che, a parità di composizione biochimica, sia la più resistente possibile, è necessario che le forze siano distribuite uniformemente in ogni elemento, e nello
stesso tempo permetta il libero passaggio di acqueo con la minima resistenza al deflusso.
Tutto questo è possibile solo con una struttura frattale, ramificata e autosomigliante; questa conserva praticamente la stessa architettura, se pur a scala minore, anche a livello del
trabecolato cribriforme, dove le sollecitazioni meccaniche delle fibre muscolari sono maggiori.
In una situazione patologica, come il glaucoma cronico, la perdita delle cellule trabecolari, con conseguente riduzione della glicoproteina della sostanza fondamentale, determinerebbe una
alterazione della composizione trabecolare. Per mantenere le stesse funzioni di distribuzione delle forze, con una riduzione della resistenza trabecolare, è necessario che si
abbia una fusione tra le trabecole adiacenti con conseguente riduzione della pervietà degli spazi intertrabecolari e un aumento della resistenza al deflusso.
In tali condizioni viene ad essere alterata la struttura autosimile frattale, realizzandosi una impalcatura anomala con grosse travate di compensazione alternate ad aree che conservano una
vaga struttura frattale.
Ancora una volta veniamo ad incontrare la struttura frattale alla base di condizioni fisiologiche, mentre la patologia si allontana dalle leggi matematiche dettate dalla natura.
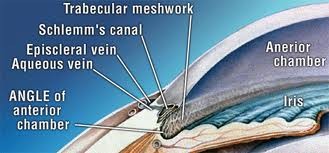
Canale di Schlemm
E' una formazione tubulare disposta attorno al limbus, a sezione ellittica.
Presenta due pareti, una esterna (sclerale) ed una interna (trabecolare): la prima è constituita da uno strato di cellule endoteliali, la seconda da due strati distinti di cellule endoteliali,
uno a contatto del trabecolato cricriforme, l'altro a contatto del canale.
Collettori post-canalicolari
Rappresentano la via di immissione dell'acqueo nel sangue e consistono in una serie di 30-40 canalicoli i quali dalla parete superficiale del canale di Schlemm si portano al plesso venoso episclerale.
Tono oculare
La pressione o tono intraoculare si mantiene su livelli relativamente costanti, suggerendo la presenza di meccanismi di regolazione per l'omeostasi del sistema.
Alcuni autori suggeriscono l'esistenza, all'interno del bulbo oculare, di barocettori. Tuttavia l'esistenza di centri e di effettori nella regolazione della pressione intraoculare è ancora
ipotetica.
Nell'ambito dei supporti nervosi, ruolo fondamentale è attribuibile al sistema autonomo, nei suoi versanti parasimpatico ed ortosimatico.
Parasimpatico
I parasimpatico-mimetici (pilocarpina) provocano la contrazione del muscolo ciliare e del muscolo sfintere dell'iride provocando un aumento del deflusso trabecolare
Ortosimpatico
Recettori beta- i recettori beta-2 sono localizzati nell'epitelio non pigmentato dei corpi ciliari e la loro stimolazione stimola la sintesi di AMPc con conseguente aumento della
produzione di acqueo. Non agiscono sul deflusso.
Recettori alfa-: i recettori alfa-1 sono localizzati a livello dei vasi, del muscolo dilatatore dell'iride e del muscolo di Muller delle palpebre. I recettori alfa-2 presentano due
localizzazioni principali: a livello presinaptico e nei corpi ciliari (NPE). La stimolazione dei primi ha un effettosimpatico-litico con diminuzione di liberazione di noradrenalina. La
stimolazione dei secondi provoca diminuzione di AMPc con diminuzione della produzione di acqueo. La stimolazione degli alfa-2 provoca inoltre un incremento del deflusso uveosclerale.(ciò è vero
peò solo per alcuni tipi di agonisti che stimolano solo la sottopopolazione alfa-2-A).
Sistema dopaminergico: è stata segnalata la presenza di recettori DA-1 a livello dei corpi ciliari. La loro stimolazione provoca aumento di produzione dell'umore acqueo. La stimolazione
dei recettori DA-2 sembrerebbe che provochi diminuzione di produzione di acqueo, agendo sulla membrana presinaptica delle fibre simpatiche periferiche.