 Ottica Campagnacci
Ottica Campagnacci










Fabrizio Campagnacci

www.otticacampagnacci.com
/home/blog

OTTICA CAMPAGNACCI aderisce alla raccolta degli occhiali usati:





OTTICA
CAMPAGNACCI
is authorized dealer of:

- Binocoli e cannocchiali
- Microscopi
- Strumenti elettronici meteo
- Altimetri e bussole
- Barometri, termometri e Galileiani
- Lenti d'ingrandimento
- Telescopi
- Visori notturni "Yukon"
- Strumenti "Suunto"
- Catalogo "C.A.I."



Apparato nervoso sensoriale

https://elvira.univr.it/bscwsci/pub/bscw.cgi/d2062609/Purves%20cap11.pdf
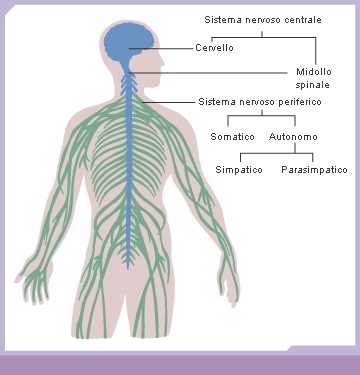
Sistema nervoso:
La percezione degli stimoli e la loro trasmissione
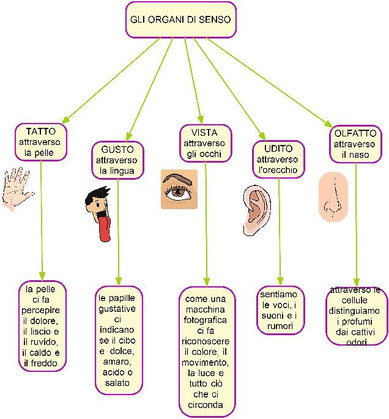
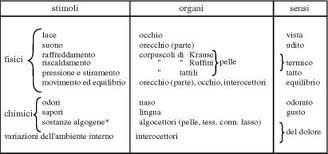
Gli esseri umani, come tutti gli animali, ricevono continuamente messaggi di vario tipo dall’ambiente in cui vivono. In base alla natura del messaggio ricevuto è possibile individuare cinque sensi: la vista, l’udito, l’olfatto, il gusto e il tatto. La sensibilità si deve alla presenza di alcuni recettori, particolari cellule capaci di captare i segnali e di reagire ai diversi stimoli. Alcune cellule sono in grado di captare i suoni, altre sono sensibili alla luce, al caldo, al freddo ecc. Esistono cinque tipi principali di recettori.
1. I termocettori, sensibili alla temperatura, trasmettono le sensazioni di caldo e freddo.
2. I nocicettori sono sensibili al dolore. Nel corpo umano sono presenti ovunque, ad eccezione del cervello. La sensazione del dolore è fondamentale per la sopravvivenza degli organismi che vengono così avvertiti della pericolosità di una lesione.
3. I meccanocettori sono sensibili a diversi tipi di pressione. Alcuni recettori meccanici particolari fanno parte degli organi dell’udito e dell’equilibrio.
4. I chemiocettori sono sensibili alla presenza di determinate sostanze chimiche. Il senso dell’olfatto e del gusto sono dovuti alla presenza di chemiocettori.
5. I recettori elettromagnetici sono sensibili all’energia legata a fenomeni quali l’elettricità o il magnetismo. Gli occhi degli animali sono dotati di fotocettori, il più comune tipo di recettori elettromagnetici, sensibili alla luce.
I recettori, posti negli organi di senso, inviano i segnali ad un complesso apparato, il sistema nervoso, che li integra, li interpreta ed elabora delle risposte. In pratica, i recettori convertono lo stimolo ricevuto in un impulso di tipo elettrico che rappresenta il modo con cui le cellule del sistema nervoso trasmettono gli stimoli.
Il sistema nervoso svolge un’azione che si articola in tre fasi:
1. L’acquisizione sensoriale è in pratica una «raccolta dei dati». Gli stimoli provenienti dagli organi di senso vengono condotti ai centri di elaborazione.
2. L’integrazione rappresenta la fase in cui le diverse informazioni vengono assemblate, interpretate ed elaborate. Queste operazioni vengono effettuate dal sistema nervoso centrale e si completano con la formulazione di una risposta allo stimolo ricevuto.
3. Lo stimolo motorio rappresenta l’impulso di risposta che parte dal centro di elaborazione e arriva alle cellule muscolari che eseguono i movimenti.Le cellule del sistema nervoso – i neuroni – sono distinguibili in tre categorie che corrispondono a ciascuna delle tre azioni appena descritte.
L’elaborazione compiuta dal sistema nervoso permette di passare dallasensazione, cioè dalla pura e semplice registrazione dello stimolo sensoriale, alla percezione, cioè ad una interpretazione del messaggio ricevuto. Le cellule nervose sono simili in tutti gli organismi. Il modo e la complessità con cui si organizzano a formare il sistema nervoso varia in relazione alle dimensioni e alle complessità dell’animale.
Alcuni organismi possiedono un sistema nervoso molto semplice, in cui le singole cellule sono associate a formare una rete nervosa priva di organi. In animali maggiormente complessi l’organizzazione del sistema nervoso possiede due caratteristiche: la centralizzazione e la cefalizzazione.
La centralizzazione è la tendenza a formare un sistema nervoso centrale separato da un sistema nervoso periferico.
La cefalizzazione è la tendenza a concentrare le strutture nervose nella zona del capo, ovvero nell’estremità anteriore rivolta verso la direzione preferenziale di movimento dell’animale.
I vertebrati possiedono un sistema nervoso altamente sviluppato, in grado di gestire il loro complesso comportamento e le molteplici funzioni specializzate.
Nel sistema nervoso si riconoscono due parti, che svolgono funzioni diverse.
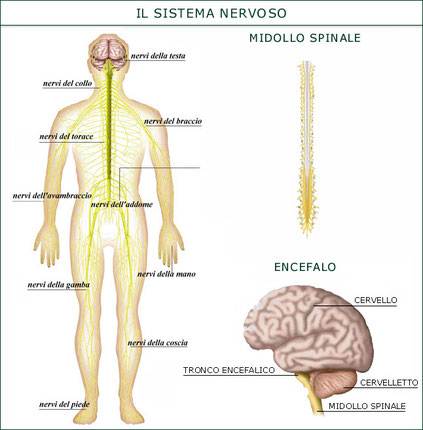
1. Il sistema nervoso centrale (SNC) costituisce il centro di elaborazione dei dati e svolge essenzialmente la funzione di integrazione, raccogliendo e interpretando gli stimoli, e fornendo le risposte. Esso è a sua volta diviso in midollo spinale ed encefalo. Il midollo spinale si trova all’interno della colonna vertebrale e si occupa dell’acquisizione sensoriale dalla pelle e dai muscoli e dell’invio dei comandi per i movimenti muscolari. L’encefalo, protetto all’interno nel cranio, è il principale organo di controllo del sistema nervoso. Esso elabora e integra tutte le informazioni provenienti dagli organi di senso e rappresenta la sede delle emozioni e dell’intelletto. Tra le diverse classi di vertebrati si registrano notevoli differenze a livello dell’encefalo. Gli uccelli e i mammiferi possiedono un encefalo che, in rapporto alle dimensioni corporee, è più voluminoso che negli altri gruppi. Lo sviluppo del cervello consente a questi animali di acquisire un gran numero di informazioni dall’ambiente circostante e di gestire complesse interazioni sociali.
2. Il sistema nervoso periferico (SNP) è costituito dalle vie di comunicazione che trasportano i messaggi dall’esterno al sistema nervoso centrale e portano le risposte dal sistema nervoso centrale all’esterno. Il sistema periferico si occupa essenzialmente dell’acquisizione sensoriale e dello stimolo motorio ed è formato dai nervi, fasci di assoni e dendriti avvolti da tessuto connettivo, e dai gangli, ammassi di corpi cellulari di neuroni. I nervi sono distinti in nervi cranici, direttamente connessi all’encefalo (come il nervo ottico), e nervi spinali, connessi al midollo spinale.
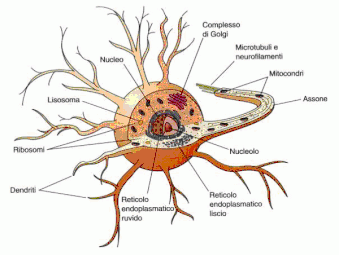
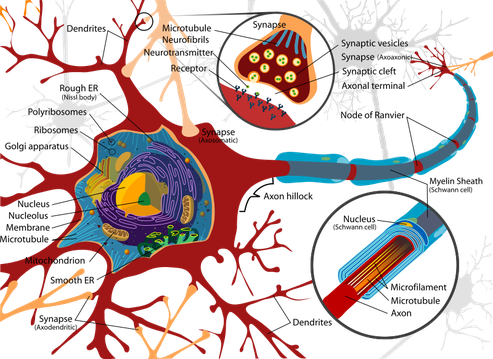
Come funziona la cellula nervosa
I neuroni funzionano in tutte le specie trasmettendo un impulso di natura elettrochimica. La membrana plasmatica di un neurone è polarizzata, cioè presenta una differenza di carica elettrica tra l’interno e l’esterno della cellula. Tale differenza è dovuta alla concentrazione degli ioni positivi (soprattutto ioni sodio Na+), maggiore all’esterno che all’interno. Questa distribuzione asimmetrica produce una differenza di potenziale elettrico detta potenziale a riposo. Il potenziale a riposo viene mantenuto dall’azione di una proteina di membrana – chiamata pompa sodio-potassio – che trasporta (in modo attivo) ioni Na+ dall’interno all’esterno della cellula e ioni potassio K+ dall’esterno verso l’interno.Gli ioni K+ possono passare liberamente attraverso delle proteine di membrana (chiamate proteine canale per il potassio) e tendono a equilibrare la loro concentrazione, spostandosi dall’interno all’esterno. Anche gli ioni Na+ si distribuirebbero uniformemente passando attraverso altre proteine (le proteine canale per il sodio), ma ciò non avviene perché quando il neurone è a riposo questi canali sono chiusi. In questo modo il potenziale a riposo è mantenuto costante, attorno al valore di circa –70 mV.
Il repentino cambiamento di questa differenza di potenziale genera l’impulso nervoso, cui segue il ripristino delle condizioni normali. Difatti, se il neurone viene stimolato, a causa dell’apertura di alcune proteine canale per il sodio il potenziale della membrana può salire da circa –70 mV a circa –50 mV, valore che è detto potenziale di soglia.
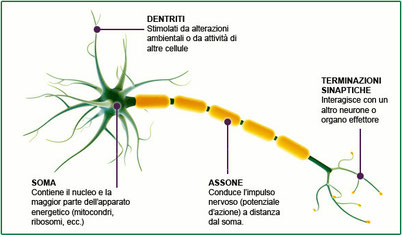
Raggiunto questo valore di soglia, molti canali per il sodio si aprono e un gran numero di ioni Na+ passano dall’esterno all’interno della cellula. Dato che la concentrazione di cariche positive all’interno aumenta, il potenziale si inverte bruscamente e raggiunge un valore di +35 mV, che viene detto potenziale d’azione. Questa sequenza di eventi è detta depolarizzazione della membrana.
Pochi istanti dopo le proteine canale per il sodio si richiudono, mentre quelle per il potassio – che nel frattempo erano chiuse – si riaprono e, grazie all’azione della pompa sodio-potassio, vengono ristabilite le condizioni a riposo. Questo processo è detto ripolarizzazione della membrana.
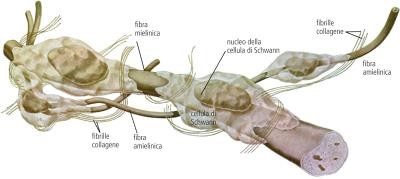
L’impulso nervoso si trasmette lungo la membrana degli assoni delle cellule nervose. Il potenziale d’azione è un fenomeno elettrochimico localizzato. Perché si verifichi la trasmissione dell’impulso, la depolarizzazione deve trasmettersi dalla zona in cui è avvenuta al tratto immediatamente adiacente. La propagazione dello stimolo avviene esclusivamente in una direzione grazie al fatto che, nel tratto interessato dal potenziale d’azione, la pompa sodio-potassio sta operando per riportare le condizioni a riposo. La concentrazione di ioni potassio nel liquido interstiziale all’esterno della cellula, per un breve istante, risulta superiore rispetto a quella delle condizioni normali a riposo; ciò determina una iperpolarizzazione della membrana che persiste finché non viene ripristinato il valore di -70 mV. In questo lasso di tempo, che dura circa 2 millisecondi, la membrana non può ricevere alcuno stimolo. Questo periodo di refrattarietà impedisce la trasmissione della depolarizzazione nella stessa direzione da cui è provenuta, evitando di fatto che la trasmissione dell’impulso si blocchi.Gli assoni delle cellule nervose umane sono avvolti dalle cellule di Schwann che formano la guaina mielinica isolante, interrotta in corrispondenza dei cosiddetti nodi di Ranvier. L’impulso nervoso procede «saltando» da un nodo di Ranvier all’altro e quindi avanza molto più rapidamente di quanto farebbe se dovesse percorrere l’intero assone.
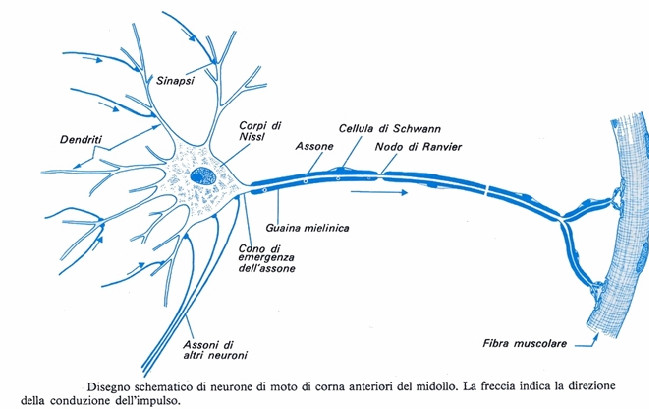

La trasmissione dell’impulso tra neuroni
I neuroni sensoriali, che acquisiscono lo stimolo sensoriale, e i neuroni di associazione, che svolgono la funzione di elaborazione e integrazione, trasmettono l’impulso ad altre cellule nervose. I neuroni motori invece passano lo stimolo alle cellule muscolari.
L’impulso nervoso viene trasferito da una cellula all’altra e alle cellule muscolari attraverso le sinapsi. Le sinapsi sono i punti di contatto tra due neuroni o tra un neurone e una cellula muscolare. Le sinapsi trasmettono alla cellula successiva l’impulso nervoso nel momento in cui esso arriva al termine dell’assone. Esistono due tipi di sinapsi: – le sinapsi elettriche,– le sinapsi chimiche.
1. Nelle sinapsi elettriche il potenziale d’azione passa da una cellula alla successiva con un meccanismo del tutto simile a quello di propagazione dell’impulso all’interno della cellula stessa. Molto spesso le sinapsi elettriche possono trasmettere l’impulso nervoso in entrambe le direzioni.
2. Le sinapsi chimiche sono presenti dove l’elaborazione degli impulsi è più varia e complessa, ad esempio nel sistema nervoso centrale. Le sinapsi chimiche presentano un piccolo spazio tra il neurone pre-sinaptico e il neurone post-sinaptico ed è questa separazione a impedire il trasferimento diretto dell’impulso elettrico. Nelle sinapsi chimiche, infatti, la trasmissione dell’impulso tra un neurone e il successivo è mediata da alcune sostanze chimiche, contenute in vescicole, dette neurotrasmettitori.
Quando l’impulso giunge al termine dell’assone, provoca l’apertura delle proteine canale degli ioni calcio Ca++ nel rigonfiamento terminale, chiamato bottone sinaptico. Gli ioni calcio sono presenti in concentrazione maggiore all’esterno della cellula e passano quindi all’interno. Questo evento induce l’esocitosi delle vescicole contenenti i neurotrasmettitori, che attraversano lo spazio sinaptico e si legano alle proteine di membrana (recettori) del neurone post-sinaptico. Il legame provoca l’apertura di canali ionici, permettendo l’entrata di ioni nel neurone post-sinaptico, provocandone la depolarizzazione e dando l’avvio all’impulso nervoso.
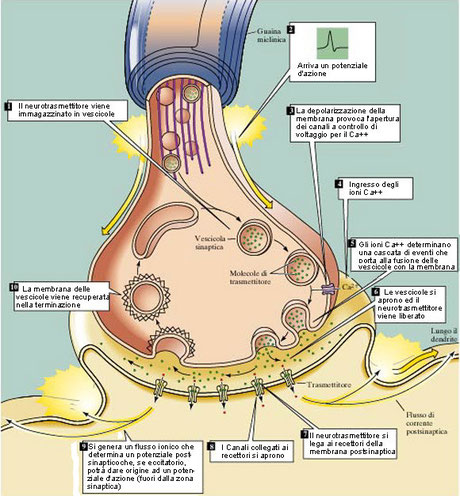
I neurotrasmettitori devono quindi essere riassorbiti e re-immagazzinati nelle vescicole, in modo che le condizioni iniziali vengano ripristinate e la cellula post-sinaptica sia pronta a ricevere un altro stimolo. Questo meccanismo permette di trasmettere l’impulso nervoso solo in un senso.
La maggior parte dei neurotrasmettitori è costituita da piccole molecole organiche quali amminoacidi o loro derivati. Alcuni neurotrasmettitori, legandosi con i recettori associati a canali ionici del Na+, ne provocano direttamente l’apertura, generando il potenziale d’azione nel neurone postsinaptico. Le sinapsi in cui questi neurotrasmettitori agiscono vengono dette eccitatorie. L’acido aspartico e l’acido glutammico sono due amminoacidi di sinapsi eccitatorie. Esistono però altri neurotrasmettitori (tra i quali l’amminoacido glicina) che agiscono a livello delle sinapsi inibitorie: legandosi con la membrana del neurone post-sinaptico, provocano l’apertura di proteine canale di tipo differente, ad esempio quelle che permettono la fuoriuscita degli ioni potassio K+. In questo caso, la depolarizzazione della membrana aumenta e rende più difficile l’avvio dell’impulso nervoso.
Alcuni neurotrasmettitori, come l’acetilcolina, sono in grado sia di trasmettere che di inibire l’impulso nervoso a seconda del tipo di recettore presente nelle cellule postsinaptiche.
L’occhio e la ricezione della luce
Nel regno animale, le poche specie completamente prive della vista sono animali che vivono in ambienti sotterranei dove la luce è assente. Gli animali che vivono alla luce possiedono organi fotorecettori di tre tipi.
L’organo fotorecettore più semplice è la macchia oculare, tipica di alcune specie di meduse e di certi vermi piatti; è costituita da un piccolo gruppo di cellule in grado di recepire la luce.Gli invertebrati quali insetti e crostacei possiedono invece un occhio composto, costituito da un gran numero di piccole unità che funzionano ciascuna come un singolo occhio. Tutti i vertebrati – per esempio gli esseri umani – e alcuni invertebrati – per esempio il calamaro – possiedono invece un occhio a lente singola, in grado di fornire un’immagine avente elevata qualità e nitidezza.
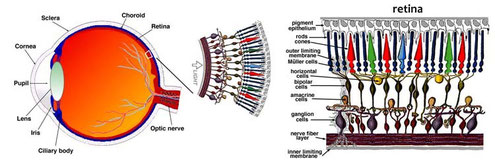
La luce entra nell’occhio dalla cornea, la parte anteriore trasparente dell’occhio. All’interno della cornea si trova l’iride, che conferisce il colore agli occhi. L’iride contiene sottili muscoli che regolano la dimensione del foro dal quale entra la luce, la pupilla. Nel caso ci si trovi in un ambiente poco luminoso, la pupilla aumenta il proprio diametro per lasciar entrare nell’occhio la massima quantità di luce possibile. Al contrario se la luce è molto forte, la pupilla si restringe diminuendo la quantità di luce che entra.
I raggi luminosi proseguono, attraversando il cristallino: la lente che mette a fuoco l’immagine sulla parete opposta del globo oculare, dove si trova la retina. Questa è formata da un tappeto di circa 130 milioni di fotorecettori che trasformano lo stimolo luminoso in impulso elettrico. La retina contiene due tipi di cellule:
– i bastoncelli, di forma allungata, sono i fotorecettori più abbondanti nell’occhio umano; sono più concentrati nella parte periferica della retina;
– i coni, cellule di forma tozza, in numero 20 volte minore dei bastoncelli, sono più numerosi nella parte centrale della retina, detta fovea.
Per mettere a fuoco un’immagine, alcuni vertebrati, come i pesci, muovono il cristallino avanti o indietro. Nell’occhio umano, invece, il cristallino cambia forma. Se l’oggetto è lontano (oltre i 6 m), il cristallino mantiene una forma allungata e focalizza i raggi luminosi paralleli sulla retina. Se l’oggetto da osservare è vicino, il cristallino aumenta la sua convessità, diventando più spesso e arrotondato, grazie alla contrazione dei muscoli che lo circondano. Questo movimento, detto accomodamento del cristallino, permette di focalizzare i raggi luminosi divergenti sulla retina. I difetti della vista più comuni sono la miopia e l’ipermetropia, che consistono in un’errata messa a fuoco delle immagini. Questi difetti sono correggibili grazie all’uso di lenti.
La ricezione degli altri stimoli
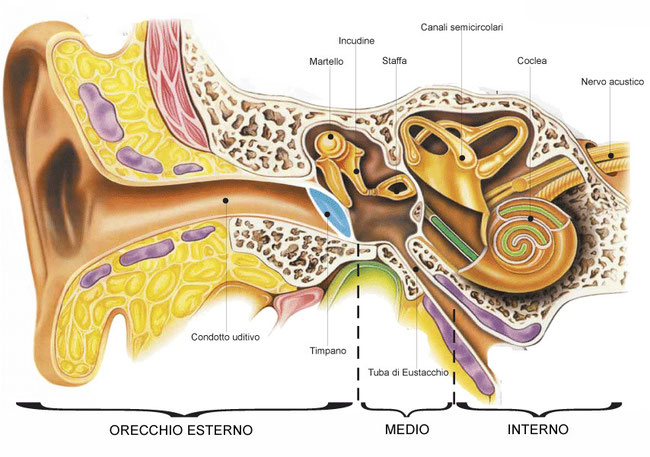
Come nella maggior parte dei vertebrati, l’orecchio umano è l’organo di senso dell’udito e contemporaneamente dell’equilibrio.L’orecchio è composto da tre parti.
1. L’orecchio esterno raccoglie i suoni e li convoglia verso l’orecchio medio. Esso è composto dal padiglione auricolare, la parte che comunemente chiamiamo orecchio, e dal meato acustico o condotto uditivo, lungo 2,5 cm, che convoglia le onde sonore contro la membrana timpanica che separa l’orecchio esterno dall’orecchio medio. Questa membrana – comunemente chiamata timpano – quando è sollecitata dalle onde sonore è in grado di vibrare e trasformare così il suono in un impulso meccanico.
2. L’orecchio medio è una piccola cavità contenente tre minuscoli ossicini disposti in sequenza: il martello, l’incudine e la staffa. La funzione dell’orecchio medio è di amplificare le vibrazioni registrate dal timpano. Tali vibrazioni vengono intensificate dalla catena di ossicini che appoggia sulla finestra ovale, una membrana che separa l’orecchio medio dall’orecchio interno.
3. L’orecchio interno, situato all’interno del cranio, contiene il labirinto, cioè l’organo dell’equilibrio, e la coclea, che rappresenta invece il vero e proprio organo dell’udito. La coclea è un lungo tubo avvolto a spirale che riceve le vibrazioni della staffa attraverso la finestra ovale. All’interno della coclea è situato l’organo del Corti che è formato da una membrana su cui sono inserite delle cellule recettrici ciliate. Le vibrazioni che la finestra ovale riceve dalla catena di ossicini producono onde di compressione nel liquido che riempie la coclea, le quali a loro volta si trasmettono alla membrana tettoria a contatto con le ciglia. La pressione trasmessa alle ciglia fa cambiare lo stato delle cellule recettrici che iniziano la trasmissione dell’impulso elettrico alle cellule del sistema nervoso collegate.
L’organo dell’equilibrio, detto labirinto membranoso, è composto da due parti – il vestibolo e i canali semicircolari – che forniscono informazioni al cervello in merito alla posizione e ai movimenti della testa. Il vestibolo è composto da due sacchi membranosi, chiamati otricolo e sacculo, ed è responsabile del mantenimento dell’equilibrio statico: ci fornisce cioè informazioni in merito all’alto e al basso (quindi alla direzione della forza di gravità) anche quando siamo immobili. I canali semicircolari sono tre, lunghi circa 12 mm ciascuno, e disposti perpendicolarmente uno all’altro come assi cartesiani. Essi sono responsabili del mantenimento dell’equilibrio dinamico, durante i movimenti angolari e rotatori della testa.
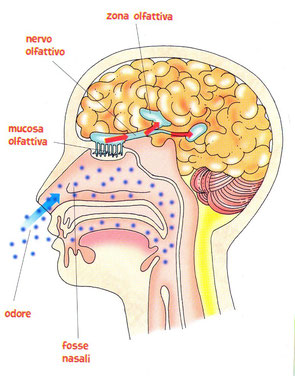
Oltre alla luce e alle onde sonore, gli esseri umani sono in grado di percepire la presenza di sostanze chimiche disperse nell’aria, attraverso l’olfatto, e all’interno dei cibi, attraverso il gusto. Questi sensi dipendono da chemiocettori: cellule capaci di captare la presenza di particolari molecole, di dare inizio alla trasduzione del segnale e di avviare l’impulso elettrico nelle cellule del sistema nervoso.
Gli esseri umani possiedono cinque milioni di recettori olfattivi (chemiocettori) e l’organo olfattivo, il naso, è capace di riconoscere ben cinquanta tipi di odori. La volta della cavità nasale è tappezzata di recettori olfattivi. L’aria inspirata è costretta nel suo percorso a fluire lungo la volta nasale; le molecole in essa contenute vengono pertanto a contatto con le cellule ciliate dei recettori che le catturano e le trattengono.
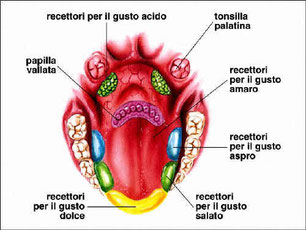
Il gusto dipende da alcuni recettori specifici che si trovano nella bocca, detti bottoni gustativi. I bottoni gustativi sono raggruppati nelle papille gustative: piccole estroflessioni della superficie della bocca, presenti in massima parte sulla lingua e, in numero molto minore, sul palato. I bottoni gustativi sono costituiti da cellule ciliate, che catturano le sostanze chimiche disciolte nella saliva grazie alle ciglia. Gli esseri umani possono distinguere quattro gusti fondamentali e ciascuno corrisponde alla stimolazione di bottoni gustativi presenti in un determinato settore della lingua. I recettori per l’amaro sono collocati nella parte posteriore della lingua e sono stimolati da particolari sostanze organiche dette alcaloidi. I recettori per l’acido sono posti nella parte mediana della lingua e vengono stimolati dagli ioni idrogeno (H+) responsabili dell’acidità. I recettori del salato sono situati soprattutto nella parte anteriore e in quella laterale della lingua e percepiscono la presenza di ioni metallici. I recettori del dolce si trovano soprattutto sulla punta della lingua e sono sollecitati dagli zuccheri e da alcuni amminoacidi.
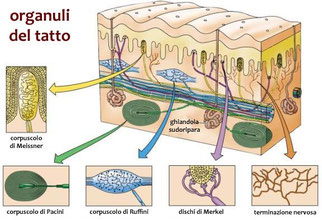
Infine, il senso del tatto si realizza grazie alla presenza di diversi tipi di meccanocettori nella pelle. Esso fornisce informazioni sulla superficie degli oggetti che tocchiamo .In ogni cm2 di pelle sono presenti mediamente 130 recettori tattili, ma in alcune zone, ad esempio sui polpastrelli, sono in numero superiore. Accanto a questi meccanocettori, nella pelle sono presenti anche numerose terminazioni nervose che svolgono la funzione di recettori del caldo, del freddo e del dolore. Ci sono recettori anche nei muscoli, nei tendini, nelle ossa e negli organi viscerali, che sono detti propriocettori. Essi ci informano costantemente sulla posizione e lo stato del corpo.
Il sistema nervoso centrale umano
Il sistema nervoso centrale umano rappresenta il sistema di elaborazione dati più complesso ed efficiente presente sul pianeta Terra ed è costituito dal midollo spinale e dall’encefalo.
Il midollo spinale si trova all’interno della colonna vertebrale e la sua funzione è quella di trasportare le informazioni dal sistema nervoso periferico verso l’encefalo e viceversa. Osservando una sezione trasversale del midollo spinale si può notare che è formato da due parti distinte:
– la sostanza grigia, situata internamente e composta principalmente dai corpi cellulari dei neuroni motori e degli interneuroni;
– la sostanza bianca, collocata all’esterno della sostanza grigia e formata da assoni e dendriti, fasci di fibre nervose che collegano i vari livelli del midollo e il midollo con l’encefalo.Al midollo spinale sono connesse le strutture del sistema nervoso periferico, come i nervi spinali e i gangli.
Il midollo spinale non si limita a trasportare informazioni ma, quando è sottoposto a stimoli intensi, può produrre risposte semplici e immediate senza l’elaborazione da parte del cervello. Questo meccanismo di difesa è detto arco riflesso semplice ed è fondamentale per limitare i danni in caso di pericolo. Nel momento in cui la terminazione di un neurone sensoriale riceve uno stimolo di dolore acuto (ad esempio se si tocca un oggetto rovente), questo si propaga lungo l’assone del neurone fino alla sostanza grigia del midollo spinale. Lo stimolo è intenso, pertanto il midollo spinale non lo conduce immediatamente al cervello per un’ulteriore elaborazione, ma lo trasferisce a un neurone motorio presente nella sostanza grigia. Il neurone motorio aziona una risposta facendo ritrarre il braccio ed evitando l’esposizione prolungata a un potenziale pericolo.
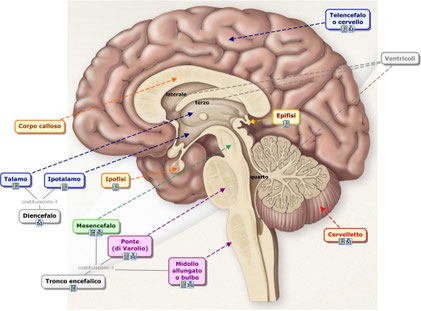
L’encefalo è l’organo di gran lunga più importante per l’acquisizione delle informazioni, per il controllo motorio e per il mantenimento dell’omeostasi nel nostro corpo.L’encefalo può essere suddiviso in quattro regioni principali.
1. Il tronco cerebrale è un’area di passaggio delle informazioni provenienti dal midollo spinale verso il resto dell’encefalo o viceversa. Il tronco contiene dei centri nervosi che controllano alcune importanti funzioni, come la respirazione e la pressione sanguigna.
2. Il cervelletto, piuttosto voluminoso, e situato alla base della scatola cranica, controlla la postura, l’equilibrio e la coordinazione dei movimenti.
3. Il diencefalo, situato al di sopra del tronco cerebrale al centro del cranio, è formato da tre strutture: talamo, ipotalamo e ipofisi. Il talamo è paragonabile a un centro di smistamento delle informazioni provenienti dagli organi di senso. L’ipotalamo controlla la temperatura corporea, il bilancio idrico e il metabolismo. Inoltre esso è sede delle emozioni, del centro del piacere e dell’assuefazione e regola anche i bioritmi giornalieri (sonno, fame). L’ipofisi, collegata all’ipotalamo tramite un sottile peduncolo, è una ghiandola fondamentale del sistema endocrino (di cui parleremo più avanti).
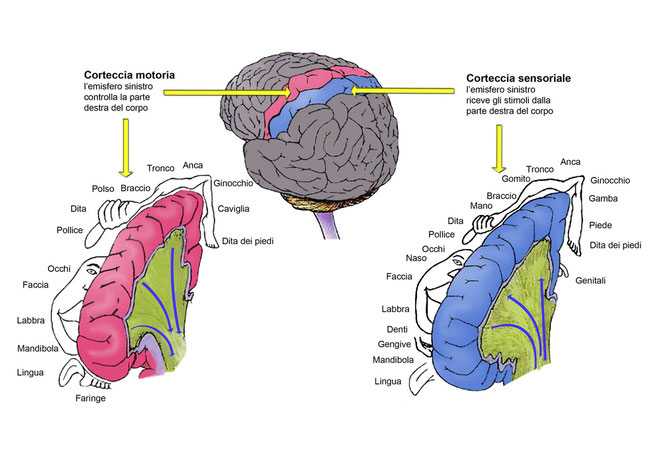
4. Il telencefalo è la parte più voluminosa di tutto l’encefalo ed è comunemente detto cervello. È costituito da sostanza bianca (all’interno) e da sostanza grigia, o corteccia cerebrale (all’esterno). Il telencefalo è costituito da due emisferi cerebrali, destro e sinistro, collegati dal corpo calloso: una spessa banda di fibre nervose che consentono una elaborazione delle informazioni comune da parte dei due emisferi. La corteccia cerebrale umana, ricca di circonvoluzioni, è formata da circa 10 miliardi di neuroni connessi da centinaia di miliardi di sinapsi ed è la sede della logica, delle capacità matematiche e linguistiche e dell’immaginazione.Il midollo spinale e l’encefalo sono circondati e protetti da tre membrane: le meningi.
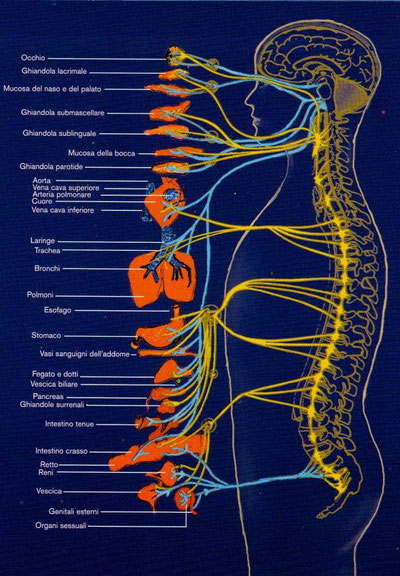
Il sistema nervoso periferico umano
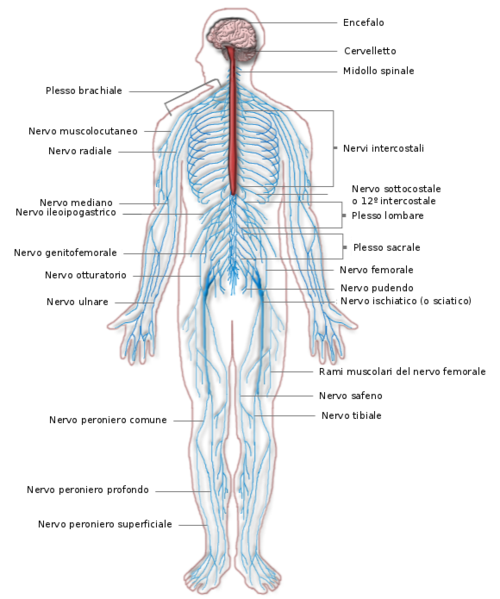
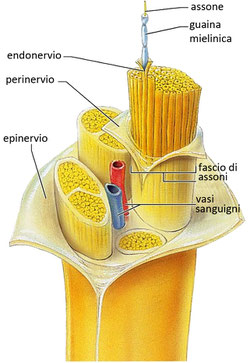
Il sistema nervoso periferico è costituito da numerosi nervi che formano una rete estesa in tutto il corpo. Ciascun nervo è formato da un gran numero di assoni appartenenti a neuroni sensoriali e neuroni motori, uniti in fasci e avvolti da tessuti connettivi che li proteggono:
- l’endonevrio avvolge un singolo assone;
- il perinevrio avvolge un fascio di assoni;
- l’epinevrio avvolge l’insieme dei fasci che formano l’intero nervo.
All’interno del fascio nervoso sono presenti anche alcuni piccoli vasi sanguigni che trasportano le sostanze nutritive e l’ossigeno. I neuroni sensoriali trasportano verso il sistema nervoso centrale le informazioni acquisite dall’ambiente esterno, attraverso gli organi di senso, e dall’ambiente interno, attraverso i recettori degli organi interni. I neuroni motori svolgono la funzione di veicolare ai muscoli e agli organi interni la risposta elaborata dal sistema nervoso centrale.
Nel sistema nervoso periferico si distinguono:
- il sistema nervoso somatico, anche detto volontario, che trasmette gli impulsi ai muscoli scheletrici;
- il sistema nervoso autonomo, anche detto involontario, che controlla le attività svolte dal corpo in modo automatico (regola il ritmo della respirazione, il battito cardiaco, i movimenti dei visceri).
Il sistema nervoso somatico è formato da un gran numero di nervi connessi con il sistema nervoso centrale e distinti in base alla posizione in due tipi:
- I nervi cranici trasportano gli impulsi da o verso l’encefalo; sono 12 paia e connettono gli organi di senso e alcune parti del capo e del viso con l’encefalo. Sono tutti nervi misti, cioè sia sensoriali sia motori, ad eccezione dei tre nervi collegati agli organi di senso (il nervo olfattivo, il nervo ottico e il nervo acustico) che sono esclusivamente nervi sensoriali.
- I nervi spinali trasportano gli impulsi da o verso il midollo spinale; sono 31 paia, sono tutti nervi misti e prendono il nome dalla regione del midollo spinale dalla quale si originano. Essi innervano tutti i muscoli e la pelle degli arti e del tronco.
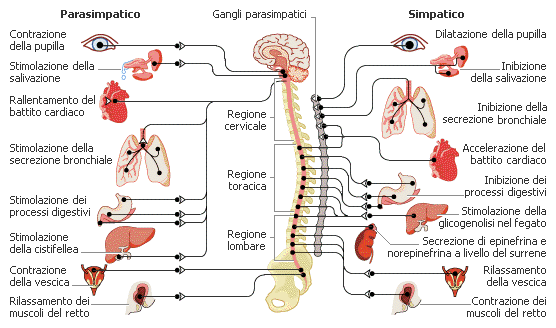
Il sistema nervoso autonomo è costituito da nervi motori che controllano, tra gli altri, il muscolo cardiaco, le ghiandole e il tessuto muscolare liscio. Il sistema nervoso autonomo è composto da due sistemi differenziati, che esercitano effetti contrapposti sugli organi che innervano:
- 1. Il sistema nervoso autonomo simpatico interviene quando l’organismo si trova in situazioni di stress, sotto sforzo e impegnato in attività intense. La sua attività causa l’accelerazione del battito cardiaco, aumenta la pressione sanguigna e la quantità di glucosio nel sangue.
- 2. Il sistema nervoso autonomo parasimpatico esercita invece un controllo esattamente opposto. La sua attività rilassa il corpo, favorisce la digestione, partecipa all’eliminazione delle feci e dell’urina e prevale nelle situazioni di tranquillità, per esempio durante il rilassamento che si ha successivamente a un pasto.
I due sistemi agiscono sugli stessi organi in maniera antagonista, cioè provocando su questi una reazione opposta.
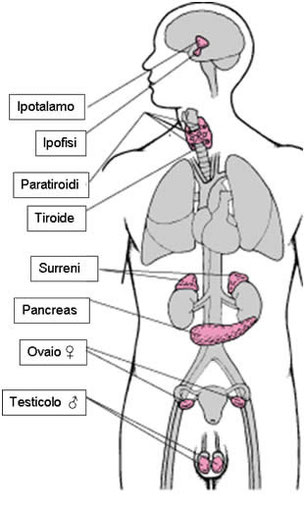
I messaggeri chimici e il sistema endocrino
Gli animali regolano molti dei loro processi vitali attraverso la produzione di alcune molecole che funzionano come messaggeri chimici. I messaggi chimici possono essere di diversa natura e agire su bersagli differenti. È possibile riconoscere tre tipi di messaggeri chimici:
- I feromoni sono sostanze, prodotte dall’organismo, che influenzano il comportamento e la fisiologia di un altro organismo della stessa specie.
- Gli ormoni sono sostanze che, una volta prodotte in organi specifici (le ghiandole endocrine), si spostano attraverso il sangue e agiscono su cellule bersaglio situate in altri organi dello stesso individuo. Esistono due tipi di ormoni: gli ormoni steroidei, derivati dalla molecola di colesterolo e solubili nei grassi, e gli ormoni proteici, solubili in acqua.
- I messaggeri paracrini sono messaggeri chimici che agiscono esclusivamente su cellule bersaglio poste nelle immediate vicinanze delle cellule da cui sono stati prodotti, senza entrare nella circolazione sanguigna.
Negli esseri umani, l’insieme delle cellule e degli organi che producono ormoni costituisce il sistema endocrino. Esso è estremamente importante in quanto regola il metabolismo, la crescita, la maturazione sessuale e numerose altre attività. Il sistema endocrino ha una funzione di regolazione simile a quella del sistema nervoso, ma agisce su tempi più lunghi. Lo stimolo nervoso si realizza in termini di frazioni di secondo, mentre un ormone raggiunge il bersaglio nell’arco di minuti, ore o anche giorni.
È costituito da numerose ghiandole, situate in diverse parti del corpo.
- Il centro di controllo di questo sistema è situato nella parte più interna dell’encefalo umano, ovvero nell’ipotalamo. Il controllo dell’ipotalamo viene esercitato attraverso la produzione di ormoni di rilasciooppure di ormoni di inibizione, i quali stimolano o bloccano l’azione di un’altra ghiandola:
- l’ipofisi. A sua volta, l’ipofisi produce ormoni che hanno come bersaglio le diverse ghiandole del corpo. L’ipofisi è costituita da due parti:
- – il lobo posteriore, chiamato neuroipofisi. La neuroipofisi secerne l’ossitocina, un ormone che nella donna provoca le contrazioni uterine durante il parto e permette la fuoriuscita del latte dalle ghiandole mammarie, e l’ormone antidiuretico, che induce la produzione di urina concentrata nei reni.
- – il lobo anteriore, chiamato adenoipofisi. L’adenoipofisi produce diversi ormoni, tra cui: la somatotropina (ormone della crescita) che induce lo sviluppo delle parti del corpo; la prolattina, che stimola la produzione di latte; le endorfine, che agiscono come antidolorifici. Controlla anche l’attività delle altre ghiandole endocrine tramite ormoni specifici.
- La ghiandola pineale (o epifisi) è un piccolo prolungamento dell’encefalo che produce la melatonina. Questo ormone influenza i ritmi biologici (sonno e veglia) e riproduttivi.
- La tiroide produce degli ormoni che regolano i processi di sviluppo dell’organismo e, nell’adulto, controllano la pressione sanguigna, il battito cardiaco e diverse funzioni del metabolismo.
- Il timo secerne un ormone che attiva lo sviluppo dei linfociti T. Questo processo avviene all’interno del timo stesso.
- Le ghiandole surrenali sono costituite da due parti: la regione interna secerne ormoni che rispondono a situazioni di stress, mentre la regione esterna produce ormoni che agiscono sull’equilibrio salino, sul livello di glucosio nel sangue e sulla risposta infiammatoria dell’organismo (come il cortisone).
- Il pancreas regola il tasso di glucosio nel sangue attraverso la produzione dell’insulina e del glucagone.
- I testicoli maschili producono ormoni androgeni (come il testosterone) che durante lo sviluppo inducono la comparsa dei caratteri sessuali maschili. Le ovaie femminili producono estrogeni e progesterone, che regolano il ciclo riproduttivo femminile e la comparsa dei caratteri sessuali femminili.
Vie ottiche:
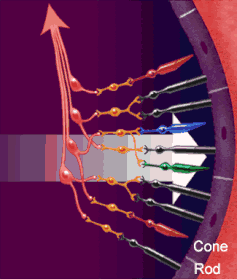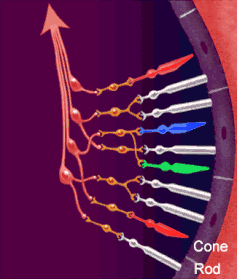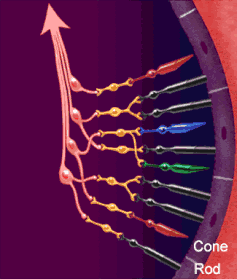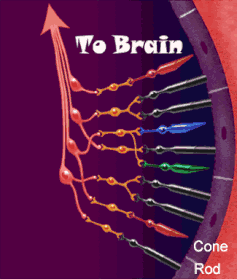
Le vie ottiche
La via ottica origina dai fotorecettori della retina (cellule dei coni per visione a colori ad alta risoluzione, cellule dei bastoncelli per visione notturna monocromatica — ma la risoluzione spaziale è molto bassa perché molti recettori convergono su un’unica fibra del nervo ottico). I fotorecettori contraggono sinapsi con le cellule bipolari della retina, che a loro volta contraggono sinapsi con le cellule multipolari della retina o cellule gangliari. Dalle cellule multipolari nascono infine le fibre ottiche che costituiscono il nervo ottico.
Vi sono tre tipi di fibre ottiche:
- Maculari: al chiasma metà fibre incrociano, l’altra metà prosegue diretta
- Nasali: si incrociano totalmente
- Temporali: non incrociano e continuano dirette
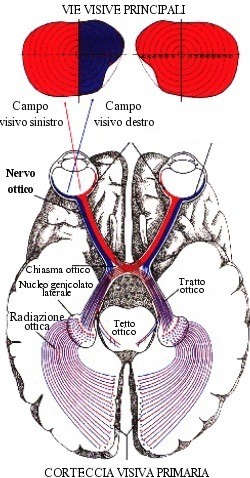
Dal chiasma ottico parte il tratto ottico, costituito da fibre maculari metà dirette e metà crociate, fibre nasali crociate, fibre temporali dirette. Il tratto ottico mette capo al corpo genicolato laterale del talamo, dove la maggior parte delle fibre termina. Dal corpo genicolato laterale del talamo originano fibre talamo-corticali che formano la radiazione ottica del Gratiolet, che passando nel braccio retrolenticolare della capsula interna e lateralmente al corno occipitale del ventricolo laterale giunge alla corteccia cerebrale dei labbri e del fondo della scissura calcarina, sede dall’area ottica primaria nel lobo occipitale.
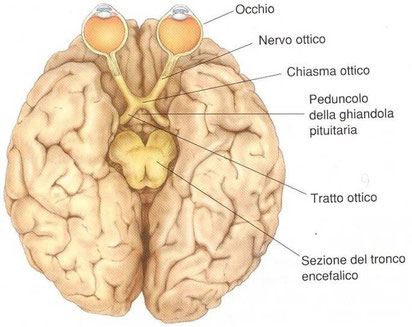
Il chiasma ottico (dal greco χίασμα, "incrocio") è la sede del parziale incrocio (decussazione) tra le fibre nervose costituenti i nervi ottici. Questi originano dagli occhi e si dirigono nell'emisfero controlaterale rispetto all'occhio d'origine.
Formazione anatomica situata nella cavità cranica e costituita dal congiungersi dei due nervi ottici. Si trova in corrispondenza della linea mediana, sopra il diaframma della sella (porzione di dura madre che ricopre la sella turcica dell’osso sfenoide su cui poggia l’ipofisi) e forma parte del pavimento del terzo ventricolo cerebrale. In corrispondenza del chiasma le fibre dei due nervi ottici si incrociano parzialmente le fibre mediali, provenienti dalla parte mediale della retina e che raccolgono le immagini della metà laterale del campo visivo, si incrociano con quelle del lato opposto: esse, insieme alle fibre della metà laterale del nervo ottico del lato opposto (che non si incrociano), formano i due bracci posteriori del chiasma, detti tratti ottici, che conducono gli impulsi visivi ai centri nervosi situati nel cervello. Dati gli stretti rapporti di vicinanza tra c. ottico, ipofisi e terzo ventricolo, diversi processi patologici propri di queste formazioni (quali tumori dell’ipofisi, meningiomi, aumento della pressione del liquor nel terzo ventricolo) possono comprimere il c. ottico determinando disturbi della visione spesso caratteristici, che vengono messi in evidenza soprattutto attraverso l’esame del campo visivo. Così se il c. ottico viene compresso nella sua parte mediana, si ha cecità delle due metà laterali del campo visivo (emianopsia bitemporale) per interruzione delle fibre provenienti dalle parti mediali delle due retine. Se il processo morboso interessa le parti laterali del c. ottico, interrompendo solo le fibre non incrociate provenienti dalla metà laterale della retina (settore temporale della retina che riceve le immagini della metà nasale del campo visivo), si avrà cecità delle due metà mediali del campo visivo (emianopsia binasale).
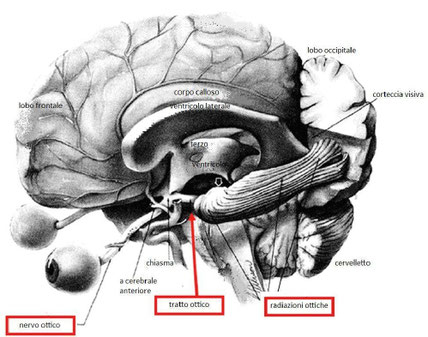
Le informazioni veicolate dai tratti ottici hanno fondamentalmente 3 destinazioni:
- Pretetto: per le risposte vegetative.
- Collicolo superiore: per essere interfacciate con le altre informazioni sensitive.
- Corpo genicolato laterale: per essere proiettate alla corteccia visiva.
Verranno ora prese in considerazione queste importanti stazioni.
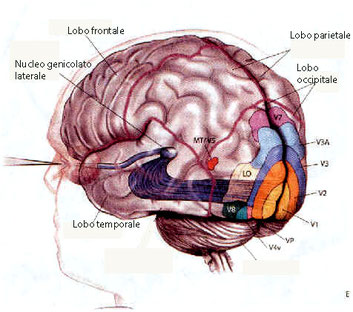
Nucleo geminato presente specularmente nella porzione destra e in quella sinistra del talamo. Il corpo genicolato laterale (CGL) è una parte del cervello preposta al trattamento dell'informazione visiva proveniente dalla retina.
È composto da due strati principali, lo strato Magnocellulare (cui fanno capo le fibre componenti la via M, adibita all'analisi delle caratteristiche di
movimento e profondità del campo visivo) e lo strato Parvocellulare (cui fanno capo le fibre componenti componenti la via P, deputate all'analisi delle forme e dei
colori).
Quindi è una stazione intermedia delle vie visive, dove le fibre dei tratti ottici entrano in contatto sinaptico con le cellule genicolate, i cui assoni si dirigono poi alla corteccia visiva primaria dello stesso lato. I campi recettivi delle cellule del corpo genicolato sono molto simili a quelli della retina: campi a simmetria circolare, con risposte di tipo on e off. Esso è diviso in 6 strati cellulari o lamine concentriche, intercalati da strati sovrapposti di assoni e dendriti; gli strati sono numerati progressivamente dalla porzione più ventrale a quella dorsale: 2 di queste hanno afferenze esclusive dalle cellule gangliari M, mentre le restanti 4 ricevono da cellule gangliari P.
In particolare:
- lamina 1: cellule M, afferenze controlaterali;
- lamina 2: cellule M, afferenze ipsilaterali;
- lamina 3: cellule P, afferenze centro-off, ipsilaterali;
- lamina 4: cellule P, afferenze centro-off, controlaterali;
- lamina 5: cellule P, afferenze centro-on, ipsilaterali;
- lamina 6: cellule P, afferenze centro-on, controlaterali.
Ciascuno strato riceve le afferenze sensitive di un solo occhio: le fibre gangliari che provengono dalla retina nasale controlaterale fanno sinapsi con le cellule degli strati 5, 3 e 2; le fibre gangliari che provengono dalla retina temporale ipsilaterale fanno sinapsi con gli strati 6, 4, 1. Le due porzioni complementari della retina di ciascun occhio inviano perciò informazioni ai singoli strati in modo topograficamente ordinato, creando in ciascuno strato una rappresentazione dell’emicampo visivo controlaterale; si produce quindi una rappresentazione completa dell’emicampo visivo controlaterale nell’intero corpo genicolato laterale, sommando le afferenze di tutti gli strati.
L'analisi in parallelo delle informazioni veicolate dalle cellule gangliari P ed M è particolarmente importante per la corretta percezione del colore e dell'illuminazione, oltre a fornire canali preferenziali per i processi di discriminazione spaziale e temporale.
| Caratteristiche dello stimolo | Sensibilità specifica cellule M | Sensibilità specifica cellule P |
|---|---|---|
| Contrasto di colore | No | Sì |
| Contrasto di luminanza | Elevato | Basso |
| Frequenza spaziale | Bassa | Elevata |
| Frequenza temporale | Elevata | Bassa |
La tabella qui sopra mostra le proprietà più importanti delle cellule M e P. Queste differenze verranno prese in considerazione successivamente per la trattazione della corteccia striata.
Nel corpo genicolato laterale, inoltre, oltre a quelle del tratto ottico, convergono molte altre fibre provenienti dalla sostanza reticolare e dalla corteccia visiva, che hanno il compito di modulare il flusso dell’informazione visiva in rapporto allo stato di veglia o di attenzione del soggetto o dell’animale. Quando la persona tende ad assopirsi o è comunque disattenta, le risposte visive sono molto attenuate, come se il cervello tendesse a ignorare le informazioni che in quel momento non ha interesse a elaborare.

Le radiazioni ottiche, dette anche radiazioni di Gratiolet, sono due estesi ventagli (uno per lato) di fibre nervose che, partite dai corpi genicolati laterali, si dirigono verso la corteccia visiva, costituendo il secondo tratto o tratto finale delle vie ottiche.
La percezione visiva è un fenomeno che dipende dalla stimolazione luminosa dei fotorecettori retinici. Tramite una reazione fotochimica si verifica una sollecitazione
dei neuroni delle vie ottiche che va a determinare la percezione visiva a livello della corteccia cerebrale.
La corteccia visiva rappresenta il punto di arrivo delle vie ottiche. Ha sede nel lobo occipitale del
cervello, il lobo più posteriore, nella zona del labbro superiore e del labbro inferiore della scissura calcarina.
Il lobo occipitale è composto da una zona posteriore, molto estesa, nella quale si presentano le fibre nervose che arrivano dalle
due fovee retiniche, la parte della retina in cui l’acuità visiva è massima, e da una zona anteriore, alquanto ridotta, nella quale si
imprimono le fibre nervose provenienti dalle restanti emiretine.
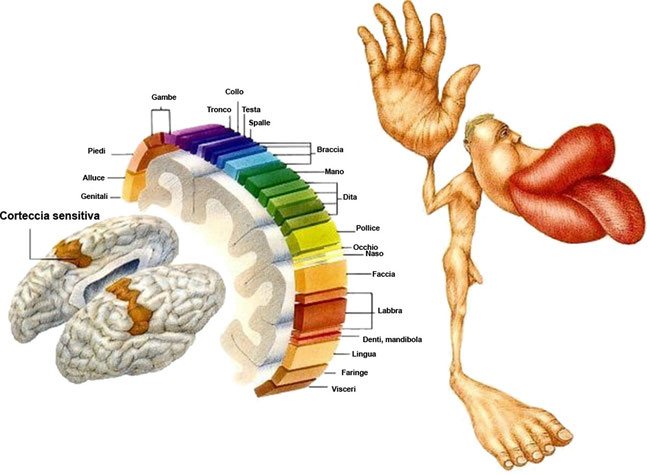
Con corteccia visiva primaria, o area 17 di Brodman, si indica la regione che si incarica dell’elaborazione delle
informazioni sugli oggetti statici e in movimento. È la prima zona in cui si dirigono le fibre che giungono dal nucleo genicolato
laterale e contiene una mappa estremamente dettagliata dell’intero campo visivo. È la prima stazione del sistema visivo in cui compaiono cellule che ricevono informazioni da
entrambi gli occhi, le cellule binoculari. L’area visiva primaria invia informazioni alle aree visive
secondarie.
La corteccia visiva secondaria, o area 18 di Brodman, e la corteccia visiva
terziaria, o area 19 di Brodman, sono definite aree associative della visione in quanto implicate nell’analisi, nel riconoscimento e nell’interpretazione delle immagini
elaborate nella corteccia visiva primaria.
La via ottica riflessa
Una parte delle fibre ottiche che non terminano nel corpo genicolato laterale del talamo raggiungono la lamina quadrigemina del mesencefalo e terminano nel tubercolo quadrigemino superiore e nel nucleo pretettale. Da qui parte la via ottica riflessa, per i movimenti riflessi in risposta a stimoli luminosi.
- Dal tubercolo quadrigemino superiore nascono fibre tetto-pontine, tetto-bulbari e tetto-spinali. Le prime due sono dirette e terminano nel ponte e nel midollo allungato; le tetto-spinali dirette si uniscono alle tetto-spinali dirette della via acustica riflessa (idem le crociate).
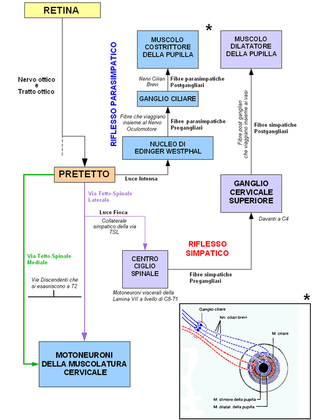
- Dal nucleo pretettale partono fibre dirette al nucleo di Edinger-Westphal del mesencefalo, dove nascono fibre per l'innervazione del muscolo costrittore della pupilla e del ciliare, responsabili rispettivamente della miosi (costrizione pupillare) in condizioni di forte luminosità e del riflesso di accomodazione visiva, ossia l'adattamento dell’occhio alla visione di oggetti vicini.
- Dal nucleo pretettale partono anche fibre dirette al nucleo intermedio laterale del corno laterale, dove ha sede il centro cilio-spinale di Budge, dal quale originano fibre per innervazione del muscolo dilatatore della pupilla per la midriasi (dilatazione pupillare) in condizioni di scarsa luminosità o stati emozionali particolari.
La zona del pretetto è formata da un gruppo di nuclei posti anteriormente e superiormente al collicolo superiore, nel punto in cui il mesencefalo si continua con il talamo. Questi nuclei sono importanti poiché sono i principali responsabili delle risposte vegetative connesse con l'apparato visivo; queste risposte sono il riflesso di costrizione e di dilatazione della pupilla, entrambi consensuali, mediati rispettivamente dal sistema parasimpatico e da sistema simpatico. Sono inoltre importarti i riflessi di puntamento in risposta ad uno stimolo visivo localizzato perifericamente al punto di fissazione foveale; tuttavia questi ultimi hanno sede solo in parte nel pretetto, poiché i movimenti oculari sono in gran parte governati da stimoli provenienti dal collicolo superiore, dalla corteccia e da alcuni nuclei che, in generale, integrano e interfacciano le diverse informazioni sensoriali, evocando una risposta di idoneo movimento oculare (si pensi ad un suono o ad uno stimolo tattile che catturano la nostra attenzione visiva). Di seguito e in figura sono trattate le importanti risposte che hanno sede nel pretetto; le risposte evocate dalla stimolazione collicolare e corticale sono trattate successivamente.

l riflesso di costrizione della pupilla è un riflesso mediato dal sistema parasimpatico molto importante indagato nella pratica clinica neurologica, utile affinché il soggetto non venga abbagliato prima che entrino in atto tutti i processi di adattamento descritti precedentemente. Una luce intensa è in grado di provocare questo riflesso innescando un processo che ha come sede principale i nuclei del pretetto. Questi proiettano assoni verso il nucleo di Edinger-Westphal (o nucleo oculomotore accessorio), un piccolo nucleo parasimpatico posto nelle immediate vicinanze del nucleo oculomotore. Il nucleo di Edinger-Westphal proietta assoni pregangliari verso il ganglio ciliare. È importante notare che queste fibre pregangliari "viaggiano insieme" al nervo oculomotore fino al ganglio ciliare. Dal ganglio ciliare si originano i nervi ciliari brevi, fibre parasimpatiche postgangliari che stimolando il muscolo costrittore della pupilla ne provocano la contrazione.
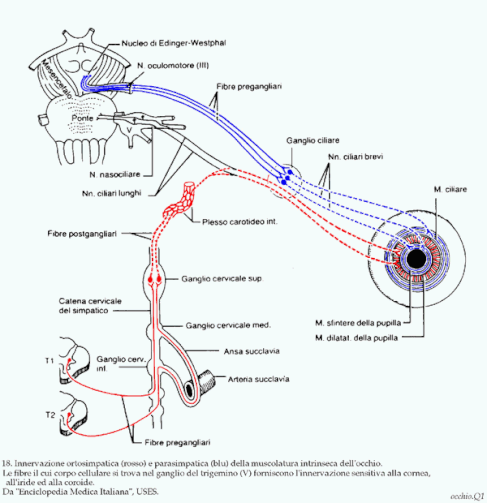
Il riflesso di dilatazione della pupilla, anch'esso indagato nella pratica neurologica, è un riflesso mediato dal sistema simpatico molto importante, utile nelle condizioni di poca luce ambientale. Permette infatti l'aumento del diametro della pupilla, consentendo alla retina di "lavorare" con un maggiore afflusso di informazioni luminose. Le poche informazioni luminose che arrivano al pretetto stimolano i neuroni di questi nuclei a contattare, tramite un collaterale della via tetto-spinale laterale, i motoneuroni viscerali della lamina VII presenti a livello di C8-T1 che vanno a costituire il centro cilio-spinale di Budge. Dal centro ciliospinale si dipartono fibre simpatiche pregangliari che risalgono i gangli paravertebrali fino a contattare sinapticamente i neuroni del ganglio cervicale superiore. Da questi si dipartono fibre simpatiche postgangliari le quali, salendo "in compagnia" delle arterie che irrorano il bulbo oculare, vanno ad innervare il muscolo dilatatore della pupilla, provocandone la contrazione e il conseguente aumento di diametro della pupilla.
Riflessi di puntamento: si immagini di stare a guardare un punto di fisso di un paesaggio. Le fovee si dispongono affinché la proiezione di quel particolare fissato cada perfettamente su di esse. L'immagine della maggior parte del paesaggio cade dunque in zone retiniche che non hanno le stesse proprietà discriminative della fovea. Tuttavia è importante, per l'uomo e per l'animale, riuscire ad avvertire nel paesaggio "di sfondo" che circonda il fuoco movimenti rapidi e improvvisi che possono comportare pericolo. "L'avvertimento della perturbazione" e analizzato dalla corteccia, dal collicolo e dal pretetto.
Istintivamente il pretetto contatta i nuclei oculomotori (poco), ma soprattutto i motoneuroni dei muscoli cervicali, sincronizzando e stimolando la loro azione in modo da portare il capo (e dunque occhi e fovee) verso il punto di perturbazione rilevato. Le vie discendenti che dal pretetto vanno a contattare questi motoneuroni sono la via tetto-spinale mediale (in larga parte) e "un po'" della via tetto-spinale laterale, utilizzata soprattutto dal riflesso di dilatazione delle pupille.
Detto questo, occorre essere cauti: benché si tratti di un vero e proprio riflesso istintivo, questo può essere soverchiato dalla corteccia (dalla volontà), nel momento in cui il soggetto decide di non rispondere ad uno stimolo che invade il suo campo visivo. Questo non succede invece con il riflessi di costrizione e dilatazione della pupilla, assolutamente "non allacciati" al controllo volontario.
Cosa importante è ricordare che i riflessi di puntamento utilizzano preferenzialmente i muscoli del collo, poiché potenti e in grado di resistere alla fatica. Non è di secondaria importanza il fatto che, ruotando il capo invece degli occhi, il soggetto si ritrova anche orecchi, naso e bocca indirizzati verso lo stimolo, elementi che possono aiutare a comprendere la natura della perturbazione.
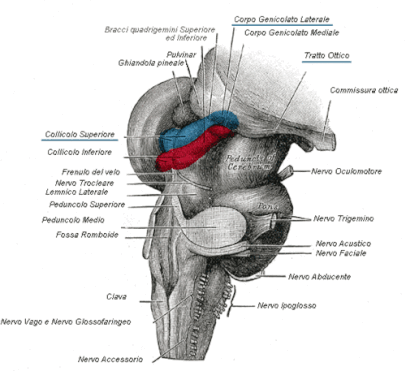
Nell'immagine qui a fianco è rappresentata la superficie posteriore del tronco dell'encefalo dopo asportazione del cervelletto. In blu è possibile apprezzare superiormente il corpo genicolato laterale e inferiormente il collicolo superiore; quest'ultimo è un importante stazione mesencefalica corrispondente al tetto ottico dei vertebrati inferiori.
L'analisi istologica rivela una complessa struttura a 7 strati, composta da diversi tipi di neuroni sui quali si scaricano informazioni provenienti dalle retine,
dai nuclei della base, dalla corteccia e dagli organi di senso. In particolare, le informazioni visive si scaricano sugli strati superficiali, le
informazioni acustiche sugli strati intermedi e le informazioni somatiche sugli strati profondi.
Analisi approfondite hanno rivelato una disposizione topografica dei neuroni del collicolo superiore; la mappa retinotopica superficiale è disposta in modo da essere congruente alla mappa
somatosensitiva e uditiva presente negli strati più profondi. Non solo: negli strati di confine sono presenti neuroni multimodali ai quali scaricano contemporaneamente
i neuroni della mappa visiva e i neuroni della mappa somatosensitava e uditiva. Il risultato è l'integrazione di diversi segnali sensoriali, fenomeno che trova riscontro nella
percezione complessa dello spazio esplorato dai sensi.
Si immagini ad esempio di vedere una rondine in volo mentre emette il caratteristico canto: per opera delle integrazioni collicolari il sistema nervoso è in grado di associare il canto (un suono)
alla rondine (l'immagine), oppure di "dirigere lo sguardo verso il suono". Non solo: grazie alla precisa mappatura degli stimoli si ha la possibilità di associare un suono ad una
data immagine e non ad un'altra. Ancora: una carezza inaspettata evocherà il puntamento istintivo della zona di cute toccata. Si può allora considerare il collicolo superiore come un insieme di
neuroni che nell'insieme formano un complesso sistema di trasduzione di coordinate.
Dato che le coordinate naturali delle tre modalità sensoriali sono differenti (coordinate retiniche per la vista, coordinate centrate sul capo per l'udito, coordinate centrate sul tronco per la
stimolazione propriocettiva), allora anche la rappresentazione spaziale degli stimoli avviene in modi differenti: le cellule visive propriamente dette codificano l'errore retinico, ovvero il
vettore che rappresenta la distanza dello stimolo dalla fovea; le cellule quasi-visive (le cellule "in transizione" tra uno stato ed un altro) codificano la posizione degli stimoli in coordinate
centrate sul capo; le cellule uditive sono sensibili sia alla posizione del suono che a quella degli occhi: sottraendo la posizione degli occhi alla posizione del suono codificata in coordinate
centrate sul capo otteniamo la posizione del suono in coordinate centrate sulla retina.
Per quanto riguarda la trattazione per il solo apparato visivo, è interessante notare come le informazioni veicolate dal tratto ottico trovino nel collicolo superiore una stazione importante non solo per il confronto sensoriale ma anche per evocare una serie di movimenti oculari strettamente connessi con l'attività dei centri superiori. Le informazioni provenienti dalla retina possono biforcarsi a livello del mesencefalo per dare una collaterale della via visiva centrale destinata al corpo genicolato laterale; inoltre esistono fibre che non si biforcano, destinate al solo (fibre tipo W) collicolo. Da qui le informazioni, dopo essere state adeguatamente integrate e confrontate, hanno diversi possibili destini:
- proiezione al pulvinar o al nucleo laterale posteriore del talamo e successivo smistamento alla corteccia striata, prestriata, temporale media e parietale;
- proiezione pontino-bulbare ipsilaterale verso i neuroni oculomotori della formazione reticolare mesencefalica e la parte rostrale della formazione reticolare pontina;
- proiezione spinale verso i motoneuroni cervicali e toracici.
Queste proiezioni testimoniano i ruolo centrale del collicolo superiore nel controllo dei movimenti oculari e cervicali, finemente regolati da centri superiori come la corteccia e i nuclei della base. In particolare, zone diverse della corteccia controllano i movimenti oculari proiettando agli strati profondi del collicolo superiore. I nuclei della base, insieme alla regione media della corteccia parietale, sono i principali responsabili dell'attenzione visiva; la regione anteriore della corteccia parietale è invece connessa con i processi di fissazione.